Histone variants for scientists
by Wajih Jawhar
Understanding the epigenome and the chromatin structure and dynamics that shape it is essential for studying epigenetics and its impact on human health and disease. Histones are fundamental constituents of chromatin that constitute a substantial regulatory layer of the epigenome, partly through chemical modifications, scientifically referred to as post-translational modifications (PTMs). These PTMs “mark” histones on specific residues and prominently on their tails that protrude from the histone octamer, which alters the structural and functional properties of chromatin as well as interactions with other regulatory factors to intricately regulate gene expression. Histones can be divided into the most abundant, “canonical” histones, and the less abundant, albeit functionally significant, “non-canonical” histones, otherwise known as histone variants.
Pairs of the four canonical histones H2A, H2B, H3 and H4 form an octameric core which DNA wraps around to form the nucleosome. These core histones are distributed across chromatin and are specifically expressed during DNA replication (replication-dependent), ensuring proper chromatin folding in daughter cells after cell division. One can imagine histones as bricks of a building that is chromatin, with DNA being the frame. Variations in the material and shape of the bricks confer specific structural properties to the part of the building where they are found. Similarly, natural variants of core histones, namely variants of histones H2A and H3, are expressed throughout the cell cycle (replication-independent) and are specifically incorporated into specific regions of chromatin to locally modify chromatin structure and function. Adding to the complexity of their contribution to epigenetic regulation, these histone variants, like their canonical counterparts, can be modified by PTMs to fine-tune their functional output.
The article will briefly introduce the well-studied human histone variants, particularly those ubiquitously expressed in cells and their proposed functions.
H3.3: The transcriptional activation histone?
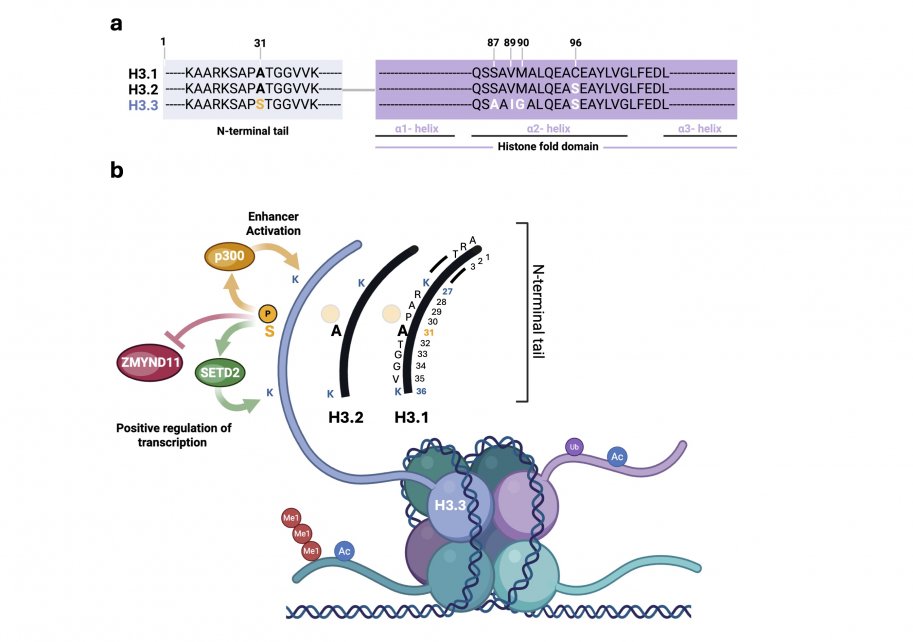
The variant Histone H3.3 has only 4 or 5 amino acid differences with canonical H3.2 and H3.1 histones, respectively. Namely residues 87, 89, 90 and 96 in the histone fold domain and residue 31 in the N terminal tail (Figure 1a). Although subtle, these unique residues are important for H3.3 histone recognition by histone chaperones and other histone readers, thereby contributing to its distinctive chromatin deposition pattern, functional roles, and involvement in specific disease mechanisms (Szenker et al., 2011; Bryant et. al, 2020 and others). For example, phosphorylation of histone H3.3 at its unique serine 31 residue promotes enhancer acetylation through stimulating p300 enzymatic activity (Martire et al., 2019). H3.3S31ph also promotes gene transcription by directly interacting with the histone methyltransferaseSETD2 to enhance its activity, and via ejecting the elongation corepressor ZMYND11 (Figure 1b). Collectively, these mechanisms are proposed to facilitate the rapid activation of stimulation-induced genes in macrophages macrophages (Wen et al., 2015; Armache et al., 2020).
H3.3 is generally found in loosely packed, transcriptionally permissive regions of chromatin, termed “euchromatin”. Unsurprisingly, it is thought to positively regulate transcription by occupying gene promoters and enhancers as well as gene bodies. H3.3 also favors deposition of active histone marks/PTMs. Notably, acetylation of the amino acid lysine (K) at positions 9, 14, 18, 23 and 27 on the histone tail, tri-methylation of K36, and di-methylation of K79 (McKittrick et al., 2004; Chow et al., 2005; Hake et al., 2006). Moreover, H3.3 containing nucleosomes exhibit a depletion in heterochromatin protein 1 (HP1) (Loyola et al., 2006). In contrast to its proposed role in reinforcing active chromatin states, H3.3 is also enriched in transcriptionally silent chromatin, such centromeres and telomeres, where it contributes to the transcriptional repression of telomeric repeats (Goldberg et al., 2010). This paradoxical distribution suggests that H3.3 can function in both active and repressive chromatin contexts. However, the precise roles of H3.3 in centromeric and telomeric chromatin organization and genome stability remain poorly understood. It is interesting to note that, this accumulation at silent chromatin could as well be due to more loading or less removal of H3.3 at these loci compared to other places in the genome (Szenker et al., 2011).
macroH2A: The transcriptional repression histone
This variant of the histone H2A is somewhat atypical relative to other variants in that it contains a significant (two-thirds) non-histone portion, called the macro domain, which protrudes out of the nucleosome (Pehrson and Fried, 1992). Initial clues on its repressive function came from a study where it was found to be concentrated in the inactive X chromosome of female mammals and implicated in the inactivation process (Costanzi and Pehrson, 1998) . Later studies confirmed and generalized its role in gene repression to other parts of the genome, including developmental genes. Indeed, macroH2A is essential for mammalian development (Buschbeck et al., 2009). Accordingly, macroH2A is also found at densely packed, transcriptionally silent chromatin, called heterochromatin, where it is important for its organization and maintenance (Douet et al., 2017).
H2AX: The DNA repair histone
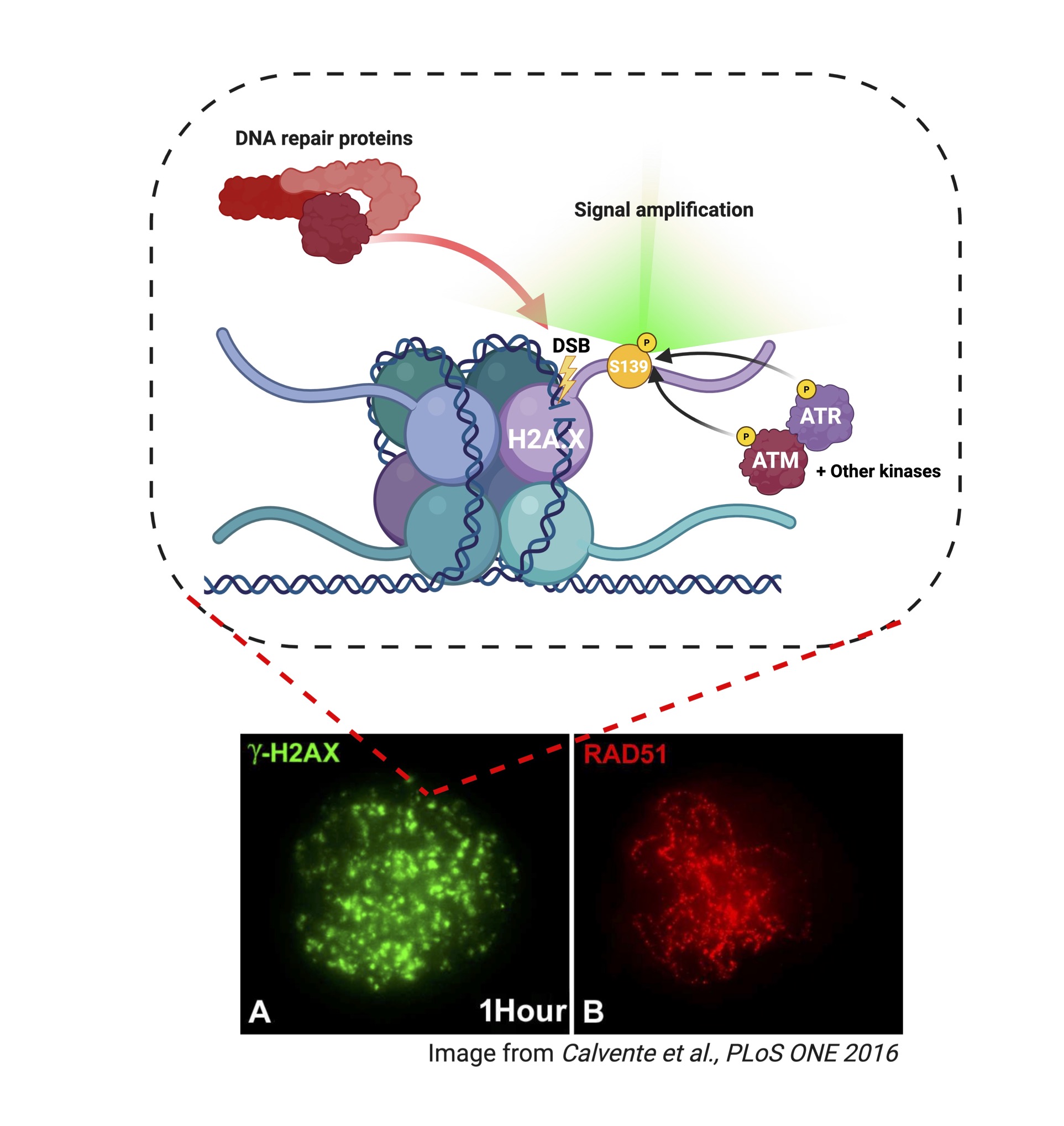
Another variant of H2A, H2AX is distributed throughout the chromatin landscape and specifically responds to one particular type of DNA damage, double strand breaks (DSBs). In response to a DSB, H2AX histones surrounding the site of the break are phosphorylated at serine 139 (S139) by several kinases of the phosphoinositide 3-kinase related kinase (PIKK) family including ATM (ataxia-telangiectasia mutated), ATR (ATM and Rad3-related) (Figure 2). Phosphorylated H2AX foci occur in the region spanning 2 megabases around the DSB. Because this was initially identified under ionizing gamma radiation that induces DSBs, phosphorylated H2AX is commonly known as ɣH2AX (Rogakou et al., 1998). The major function of ɣH2AX is acting as a signal for recruitment of DNA repair factors, ensuring genome stability (Hauer and Gasser, 2017).
H2AZ: The multifunctional histone
This H2A variant has long puzzled researchers by its involvement in very diverse, sometimes opposing processes. H2AZ has been described to play important roles in genome maintenance and expression such as heterochromatin regulation, proper chromosome segregation, DNA repair, and transcription (Rangasamy et al, 2003 & 2004; Adam et al, 2001). Paradoxically, it has been shown to positively regulate both transcriptional activation and repression. Studies characterizing its PTMs have, at least in part, begun to resolve the enigma. Apart from SUMOylation which has been linked to DNA repair, H2AZ modifications have been most heavily investigated in the context of transcriptional regulation. To regulate transcription, H2AZ is enriched at promoters and enhancers. There is strong evidence for acetylation of K4, K7 and K11 inducing/positively regulating transcription via nucleosome instability and enhanced transcription factor binding. On the other hand, mono-ubiquitination of K120, K121 and K125 is associated with gene repression. Additionally, methylation functions on opposite ends, with K7me negatively correlating while K101me2 positively correlating with transcription (Giaimo et al., 2019)
CENP-A: the chromosome segregation histone
The other well-described variant of H3, CENP-A plays a specialized role in chromosome segregation during mitosis, where it is specifically found at chromosome centromeres, allowing the binding of kinetochore proteins and attachment of sister chromatids to opposite ends of the microtubule spindle. CENP-A is therefore essential for cell division. Interestingly, centromere identity is largely defined by CENP-A positioning rather than by the underlying DNA sequence. (Gambogi and Black, 2019).
Histone variants in Cancer
Lastly, abnormal changes in the DNA code can instruct the cell to produce pathogenic variants of histones. Due to the growing body of evidence on their role in cancer, they are commonly referred to as “oncohistones”. You can read more about the different oncohistones, specifically commonly investigated oncohistones, and their role in cancer and other disease here!
References:
Szenker E, Ray-Gallet D, Almouzni G. The double face of the histone variant H3.3. Cell Res. 2011 Mar;21(3):421-34. doi: 10.1038/cr.2011.14. Epub 2011 Jan 25. PMID: 21263457; PMCID: PMC3193428.
Bryant L, Li D, Cox SG, Marchione D, et al., Histone H3.3 beyond cancer: Germline mutations in Histone 3 Family 3A and 3B cause a previously unidentified neurodegenerative disorder in 46 patients. Sci Adv. 2020 Dec 2;6(49):eabc9207. doi: 10.1126/sciadv.abc9207. PMID: 33268356; PMCID: PMC7821880.
Martire S, Gogate AA, Whitmill A, et al., Phosphorylation of histone H3.3 at serine 31 promotes p300 activity and enhancer acetylation. Nat Genet. 2019 Jun;51(6):941-946. doi: 10.1038/s41588-019-0428-5. Epub 2019 May 31. PMID: 31152160; PMCID: PMC6598431.
Wen H, Li Y, Xi Y, et al., ZMYND11 links histone H3.3K36me3 to transcription elongation and tumour suppression. Nature. 2014 Apr 10;508(7495):263-8. doi: 10.1038/nature13045. Epub 2014 Mar 2. PMID: 24590075; PMCID: PMC4142212.
Armache A, Yang S, Martínez de Paz A et al., Histone H3.3 phosphorylation amplifies stimulation-induced transcription. Nature. 2020 Jul;583(7818):852-857. doi: 10.1038/s41586-020-2533-0. Epub 2020 Jul 22. PMID: 32699416; PMCID: PMC7517595.
McKittrick E, Gafken PR, Ahmad K, Henikoff S. Histone H3.3 is enriched in covalent modifications associated with active chromatin. Proc Natl Acad Sci U S A. 2004 Feb 10;101(6):1525-30. doi: 10.1073/pnas.0308092100. Epub 2004 Jan 19. PMID: 14732680; PMCID: PMC341768.
Chow CM, Georgiou A, Szutorisz H, et al., EMBO Rep. 2005 Apr;6(4):354-60. doi: 10.1038/sj.embor.7400366. PMID: 15776021; PMCID: PMC1299280.
Hake SB, Allis CD. Histone H3 variants and their potential role in indexing mammalian genomes: the "H3 barcode hypothesis". Proc Natl Acad Sci U S A. 2006 Apr 25;103(17):6428-35. doi: 10.1073/pnas.0600803103. Epub 2006 Mar 29. PMID: 16571659; PMCID: PMC1564199.
Loyola A, Bonaldi T, Roche D, Imhof A, Almouzni G. PTMs on H3 variants before chromatin assembly potentiate their final epigenetic state. Mol Cell. 2006 Oct 20;24(2):309-16. doi: 10.1016/j.molcel.2006.08.019. PMID: 17052464.
Goldberg AD, Banaszynski LA, Noh Kmet al., Distinct factors control histone variant H3.3 localization at specific genomic regions. Cell. 2010 Mar 5;140(5):678-91. doi: 10.1016/j.cell.2010.01.003. PMID: 20211137; PMCID: PMC2885838.
Pehrson JR, Fried VA. MacroH2A, a core histone containing a large nonhistone region. Science. 1992 Sep 4;257(5075):1398-400. doi: 10.1126/science.1529340. PMID: 1529340.
Costanzi C, Pehrson JR. Histone macroH2A1 is concentrated in the inactive X chromosome of female mammals. Nature. 1998 Jun 11;393(6685):599-601. doi: 10.1038/31275. PMID: 9634239.
Buschbeck M, Uribesalgo I, Wibowo I, et al., The histone variant macroH2A is an epigenetic regulator of key developmental genes. Nat Struct Mol Biol. 2009 Oct;16(10):1074-9. doi: 10.1038/nsmb.1665. Epub 2009 Sep 6. PMID: 19734898.
Douet J, Corujo D, Malinverni R, et al., MacroH2A histone variants maintain nuclear organization and heterochromatin architecture. J Cell Sci. 2017 May 1;130(9):1570-1582. doi: 10.1242/jcs.199216. Epub 2017 Mar 10. PMID: 28283545.
Rogakou EP, Pilch DR, Orr AH, Ivanova VS, Bonner WM. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J Biol Chem. 1998 Mar 6;273(10):5858-68. doi: 10.1074/jbc.273.10.5858. PMID: 9488723.
Hauer MH, Gasser SM. Chromatin and nucleosome dynamics in DNA damage and repair. Genes Dev. 2017 Nov 15;31(22):2204-2221. doi: 10.1101/gad.307702.117. PMID: 29284710; PMCID: PMC5769766.
Rangasamy D, Berven L, Ridgway P, Tremethick DJ. Pericentric heterochromatin becomes enriched with H2A.Z during early mammalian development. EMBO J. 2003;22(7):1599–1607. doi: 10.1093/emboj/cdg160.
Rangasamy D, Greaves I, Tremethick DJ. RNA interference demonstrates a novel role for H2AZ in chromosome segregation. Nat Struct Mol Biol. 2004;11(7):650–655. doi: 10.1038/nsmb786.
Adam M, Robert F, Larochelle M, Gaudreau L. H2A.Z is required for global chromatin integrity and for recruitment of RNA polymerase II under specific conditions. Mol Cell Biol. 2001;21(18):6270–6279. doi: 10.1128/MCB.21.18.6270-6279.2001
Giaimo BD, Ferrante F, Herchenröther A, Hake SB, Borggrefe T. The histone variant H2A.Z in gene regulation. Epigenetics Chromatin. 2019 Jun 14;12(1):37. doi: 10.1186/s13072-019-0274-9. PMID: 31200754; PMCID: PMC6570943.
Craig W. Gambogi, Ben E. Black; The nucleosomes that mark centromere location on chromosomes old and new. Essays Biochem 23 April 2019; 63 (1): 15–27. doi: https://doi.org/10.1042/EBC20180060