Les variantes d'histones pour les scientifiques
par Wajih Jawhar
La compréhension de l'épigénome, ainsi que de la structure et de la dynamique de la chromatine qui le façonnent, est essentielle pour étudier l'épigénétique et son impact sur la santé et les maladies humaines. Les histones sont des composants essentiels de la chromatine et représentent une couche majeure de régulation de l’épigénome, notamment grâce à des modifications chimiques appelées les modifications post-traductionnelles (MPT). Ces MPT marquent des résidus spécifiques des histones, en particulier au niveau de leurs queues N-terminales qui dépassent de l’octamère, entraînant des changements dans la structure et la fonction de la chromatine. Elles influencent également les interactions avec divers facteurs régulateurs, contribuant ainsi à une régulation fine et complexe de l’expression génique. Les histones se divisent en deux grandes catégories : les histones canoniques, les plus abondantes, et les histones non canoniques, moins abondantes mais tout aussi importantes sur le plan fonctionnel, également appelées variants d’histones.
Les quatre histones canoniques H2A, H2B, H3 et H4 s’assemblent en paires pour former un octamère autour duquel l’ADN s’enroule, constituant ainsi le nucléosome. Ces histones sont largement distribuées dans la chromatine et leur expression est étroitement liée à la réplication de l’ADN, assurant un repliement adéquat de la chromatine dans les cellules filles après la division cellulaire. On peut les comparer à des briques formant la structure d’un bâtiment, la chromatine, tandis que l’ADN en représente l’armature. Les variations dans la composition ou la forme de ces briques confèrent des propriétés structurelles spécifiques aux différentes régions du bâtiment (chromatine).
Dans ce contexte, les variants des histones du noyau, principalement issus des familles H2A et H3, sont exprimés tout au long du cycle cellulaire, indépendamment de la réplication. Ils sont incorporés de manière ciblée dans certaines régions de la chromatine, où ils modulent localement sa structure et sa fonction. Comme les histones canoniques, ces variants peuvent également subir des MPT, ce qui renforce encore la complexité de leur rôle dans la régulation épigénétique.
Cet présente brièvement les principaux variants d’histones humains bien caractérisés, en mettant l’accent sur ceux exprimés de manière ubiquitaire, ainsi que sur leurs fonctions supposées.
H3.3 : l'histone responsable de l'activation transcriptionnelle ?
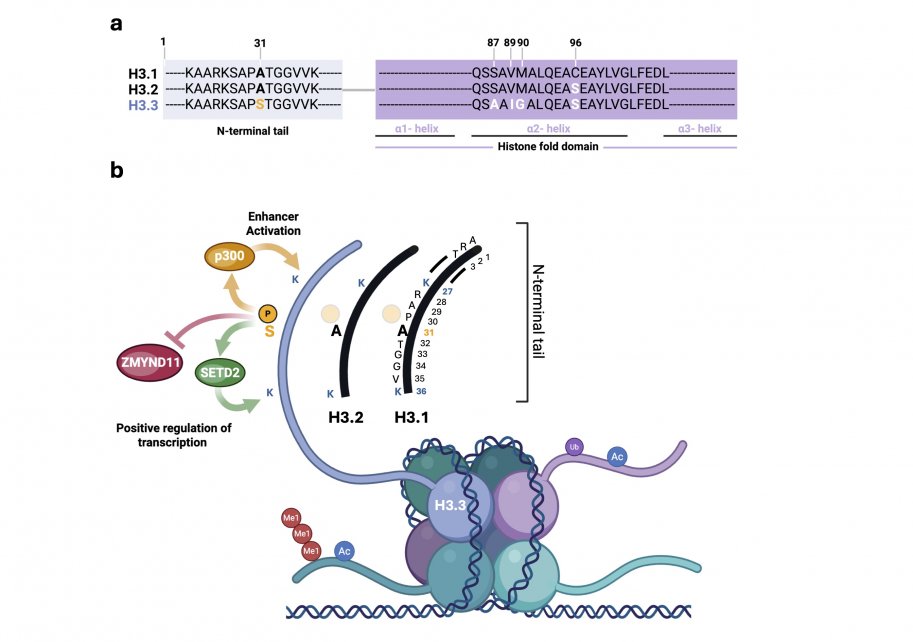
La variante d’histone H3.3 ne diffère des histones canoniques H3.1 et H3.2 que par 4 à 5 acides aminés. Ces différences concernent les résidus 87, 89, 90 et 96 au niveau du domaine globulaire de repliement, ainsi que le résidu 31 situé dans la queue N-terminale (figure 1a). Bien que minimes, ces variations sont déterminantes pour la reconnaissance spécifique de H3.3 par les chaperons d’histones et d’autres protéines « lecteurs ». Elles contribuent ainsi à son mode de dépôt particulier dans la chromatine, à ses fonctions spécifiques et à son implication dans certains processus pathologiques (Szenker et al., 2011 ; Bryant et al., 2020, entre autres).
Par exemple, la phosphorylation de H3.3 sur son résidu unique sérine 31 favorise l’acétylation des enhancers en stimulant l’activité de l’acétyltransférase p300 (Martire et al., 2019). Cette modification (H3.3S31ph) participe également à l’activation transcriptionnelle en interagissant directement avec la méthyltransférase SETD2, dont elle renforce l’activité, tout en facilitant l’éviction du corépresseur d’élongation ZMYND11 (figure 1b).
Dans l’ensemble, ces mécanismes contribueraient à une activation rapide des gènes en réponse à une stimulation, notamment dans les macrophages (Wen et al., 2015 ; Armache et al., 2020).
Le variant H3.3 est principalement localisée dans des régions de chromatine peu compactées et transcriptionnellement actives, appelées euchromatine. Elle est généralement associée à une régulation positive de la transcription, en étant enrichie au niveau des promoteurs, des enhancers ainsi que des corps des gènes. H3.3 est également liée au dépôt de marques histones actives, notamment l’acétylation des lysines (K) en positions 9, 14, 18, 23 et 27 sur la queue de l’histone, ainsi que la triméthylation de K36 et la diméthylation de K79 (McKittrick et al., 2004 ; Chow et al., 2005 ; Hake et al., 2006). Par ailleurs, les nucléosomes contenant H3.3 sont caractérisés par une réduction de l’association avec la protéine HP1, un marqueur classique de l’hétérochromatine, c’est-à-dire une forme de chromatine compacte et transcriptionnellement inactive (Loyola et al., 2006). Cependant, et de manière plus surprenante, H3.3 est également enrichie dans certaines régions transcriptionnellement silencieuses, comme les centromères et les télomères, où elle participerait à la répression des séquences répétées télomériques (Goldberg et al., 2010). Cette distribution apparemment paradoxale suggère que H3.3 peut intervenir à la fois dans des contextes de chromatine active et répressive. Néanmoins, ses rôles précis dans l’organisation de la chromatine centromérique et télomérique, ainsi que dans le maintien de la stabilité du génome, restent encore mal définis. Il est d’ailleurs possible que son accumulation dans ces régions résulte soit d’un recrutement accru, soit d’une élimination plus limitée de H3.3 à ces loci comparativement au reste du génome (Szenker et al., 2011).

macroH2A : l'histone de répression transcriptionnelle
Cette variante de l’histone H2A se distingue des autres par sa structure atypique, caractérisée par la présence d’un large domaine non histonique, représentant environ les deux tiers de la protéine, appelé « macro-domaine », qui s’étend en dehors du nucléosome (Pehrson et Fried, 1992). Les premiers éléments suggérant une fonction répressive de cette variante proviennent d’une étude montrant son enrichissement sur le chromosome X inactif chez les mammifères femelles, ainsi que son implication dans le processus d’inactivation (Costanzi et Pehrson, 1998). Des travaux ultérieurs ont confirmé et élargi ce rôle à d’autres régions du génome, notamment à des gènes impliqués dans le développement. Ainsi, la macroH2A apparaît comme un acteur clé du développement chez les mammifères (Buschbeck et al., 2009). En cohérence avec cette fonction, elle est principalement associée à des régions de chromatine fortement compactées et transcriptionnellement silencieuses (hétérochromatine), où elle contribue à leur organisation et à leur maintien (Douet et al., 2017).
H2AX : l'histone impliquée dans la réparation de l'ADN
Une autre variante de H2A, H2AX, est largement distribuée dans l’ensemble de la chromatine et joue un rôle clé dans la réponse aux dommages de l’ADN, en particulier aux cassures double brin (DSB). En réponse à ce type de lésion, les histones H2AX situées autour du site de la cassure sont phosphorylées sur la sérine 139 (S139) par plusieurs kinases de la famille des phosphoinositide 3-kinases apparentées (PIKK), notamment ATM (ataxia-telangiectasia mutated) et ATR (ATM and Rad3-related) (figure 2). Cette phosphorylation s’étend sur une large région chromatinienne pouvant atteindre environ 2 mégabases autour de la cassure, formant des foyers détectables de H2AX phosphorylée. Initialement décrite dans des cellules exposées à des rayonnements ionisants induisant des DSB, cette forme phosphorylée est communément appelée γH2AX (Rogakou et al., 1998). La γH2AX agit principalement comme un signal de recrutement pour les protéines impliquées dans la réparation de l’ADN, contribuant ainsi au maintien de la stabilité du génome (Hauer et Gasser, 2017).

H2AZ : l'histone multifonctionnelle
Cette variante de l’histone H2A suscite un grand intérêt en raison de son implication dans des processus divers, parfois apparemment contradictoires. H2A.Z joue un rôle crucial dans le maintien et l’expression du génome, notamment en participant à la régulation de l’hétérochromatine, à la ségrégation chromosomique, à la réparation de l’ADN et à la transcription (Rangasamy et al., 2003 & 2004 ; Adam et al., 2001). Paradoxalement, H2A.Z est impliquée à la fois dans l’activation et la répression transcriptionnelles. L’étude de ses MPT a permis de mieux comprendre cette dualité. Outre la SUMOylation, associée à la réparation de l’ADN, les MPT de H2A.Z ont été particulièrement étudiées dans le contexte de la régulation transcriptionnelle. Pour moduler la transcription, H2A.Z s’accumule au niveau des promoteurs et des enhancers. L’acétylation des lysines K4, K7 et K11 favorise l’activation transcriptionnelle, en rendant les nucléosomes plus instables et en facilitant la liaison des facteurs de transcription. À l’inverse, la mono-ubiquitination des lysines K120, K121 et K125 est associée à la répression génique. Par ailleurs, la méthylation agit différemment selon le site : K7me est corrélée négativement à la transcription, tandis que K101me2 présente une corrélation positive (Giaimo et al., 2019).
CENP-A : l'histone de la ségrégation chromosomique
Une autre variante bien caractérisée de l’histone H3, CENP-A, joue un rôle spécifique dans la ségrégation chromosomique lors de la mitose. Elle se localise exclusivement au niveau des centromères, où elle permet la liaison des protéines du kinétochore et l’attachement des chromatides sœurs aux pôles opposés du fuseau microtubulaire. CENP-A est donc indispensable à la division cellulaire. Il est intéressant de noter que l’identité du centromère est principalement déterminée par le positionnement de CENP-A plutôt que par la séquence d’ADN sous-jacente (Gambogi et Black, 2019).
Les variants d'histones dans le cancer
Enfin, des altérations anormales du code génétique peuvent conduire à l’expression de variants pathogènes d’histones, communément appelés « oncohistones ». Ces dernières années, un nombre croissant de preuves a mis en évidence leur implication dans le cancer, ainsi que dans d’autres pathologies, ce qui explique l’intérêt croissant et soutenu pour ces protéines. Des informations plus détaillées sur les différentes oncohistones et leur rôle dans le cancer et d’autres maladies sont disponibles ici!
References:
Szenker E, Ray-Gallet D, Almouzni G. The double face of the histone variant H3.3. Cell Res. 2011 Mar;21(3):421-34. doi: 10.1038/cr.2011.14. Epub 2011 Jan 25. PMID: 21263457; PMCID: PMC3193428.
Bryant L, Li D, Cox SG, Marchione D, et al., Histone H3.3 beyond cancer: Germline mutations in Histone 3 Family 3A and 3B cause a previously unidentified neurodegenerative disorder in 46 patients. Sci Adv. 2020 Dec 2;6(49):eabc9207. doi: 10.1126/sciadv.abc9207. PMID: 33268356; PMCID: PMC7821880.
Martire S, Gogate AA, Whitmill A, et al., Phosphorylation of histone H3.3 at serine 31 promotes p300 activity and enhancer acetylation. Nat Genet. 2019 Jun;51(6):941-946. doi: 10.1038/s41588-019-0428-5. Epub 2019 May 31. PMID: 31152160; PMCID: PMC6598431.
Wen H, Li Y, Xi Y, et al., ZMYND11 links histone H3.3K36me3 to transcription elongation and tumour suppression. Nature. 2014 Apr 10;508(7495):263-8. doi: 10.1038/nature13045. Epub 2014 Mar 2. PMID: 24590075; PMCID: PMC4142212.
Armache A, Yang S, Martínez de Paz A et al., Histone H3.3 phosphorylation amplifies stimulation-induced transcription. Nature. 2020 Jul;583(7818):852-857. doi: 10.1038/s41586-020-2533-0. Epub 2020 Jul 22. PMID: 32699416; PMCID: PMC7517595.
McKittrick E, Gafken PR, Ahmad K, Henikoff S. Histone H3.3 is enriched in covalent modifications associated with active chromatin. Proc Natl Acad Sci U S A. 2004 Feb 10;101(6):1525-30. doi: 10.1073/pnas.0308092100. Epub 2004 Jan 19. PMID: 14732680; PMCID: PMC341768.
Chow CM, Georgiou A, Szutorisz H, et al., EMBO Rep. 2005 Apr;6(4):354-60. doi: 10.1038/sj.embor.7400366. PMID: 15776021; PMCID: PMC1299280.
Hake SB, Allis CD. Histone H3 variants and their potential role in indexing mammalian genomes: the "H3 barcode hypothesis". Proc Natl Acad Sci U S A. 2006 Apr 25;103(17):6428-35. doi: 10.1073/pnas.0600803103. Epub 2006 Mar 29. PMID: 16571659; PMCID: PMC1564199.
Loyola A, Bonaldi T, Roche D, Imhof A, Almouzni G. PTMs on H3 variants before chromatin assembly potentiate their final epigenetic state. Mol Cell. 2006 Oct 20;24(2):309-16. doi: 10.1016/j.molcel.2006.08.019. PMID: 17052464.
Goldberg AD, Banaszynski LA, Noh Kmet al., Distinct factors control histone variant H3.3 localization at specific genomic regions. Cell. 2010 Mar 5;140(5):678-91. doi: 10.1016/j.cell.2010.01.003. PMID: 20211137; PMCID: PMC2885838.
Pehrson JR, Fried VA. MacroH2A, a core histone containing a large nonhistone region. Science. 1992 Sep 4;257(5075):1398-400. doi: 10.1126/science.1529340. PMID: 1529340.
Costanzi C, Pehrson JR. Histone macroH2A1 is concentrated in the inactive X chromosome of female mammals. Nature. 1998 Jun 11;393(6685):599-601. doi: 10.1038/31275. PMID: 9634239.
Buschbeck M, Uribesalgo I, Wibowo I, et al., The histone variant macroH2A is an epigenetic regulator of key developmental genes. Nat Struct Mol Biol. 2009 Oct;16(10):1074-9. doi: 10.1038/nsmb.1665. Epub 2009 Sep 6. PMID: 19734898.
Douet J, Corujo D, Malinverni R, et al., MacroH2A histone variants maintain nuclear organization and heterochromatin architecture. J Cell Sci. 2017 May 1;130(9):1570-1582. doi: 10.1242/jcs.199216. Epub 2017 Mar 10. PMID: 28283545.
Rogakou EP, Pilch DR, Orr AH, Ivanova VS, Bonner WM. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J Biol Chem. 1998 Mar 6;273(10):5858-68. doi: 10.1074/jbc.273.10.5858. PMID: 9488723.
Hauer MH, Gasser SM. Chromatin and nucleosome dynamics in DNA damage and repair. Genes Dev. 2017 Nov 15;31(22):2204-2221. doi: 10.1101/gad.307702.117. PMID: 29284710; PMCID: PMC5769766.
Rangasamy D, Berven L, Ridgway P, Tremethick DJ. Pericentric heterochromatin becomes enriched with H2A.Z during early mammalian development. EMBO J. 2003;22(7):1599–1607. doi: 10.1093/emboj/cdg160.
Rangasamy D, Greaves I, Tremethick DJ. RNA interference demonstrates a novel role for H2AZ in chromosome segregation. Nat Struct Mol Biol. 2004;11(7):650–655. doi: 10.1038/nsmb786.
Adam M, Robert F, Larochelle M, Gaudreau L. H2A.Z is required for global chromatin integrity and for recruitment of RNA polymerase II under specific conditions. Mol Cell Biol. 2001;21(18):6270–6279. doi: 10.1128/MCB.21.18.6270-6279.2001
Giaimo BD, Ferrante F, Herchenröther A, Hake SB, Borggrefe T. The histone variant H2A.Z in gene regulation. Epigenetics Chromatin. 2019 Jun 14;12(1):37. doi: 10.1186/s13072-019-0274-9. PMID: 31200754; PMCID: PMC6570943.
Craig W. Gambogi, Ben E. Black; The nucleosomes that mark centromere location on chromosomes old and new. Essays Biochem 23 April 2019; 63 (1): 15–27. doi: https://doi.org/10.1042/EBC20180060